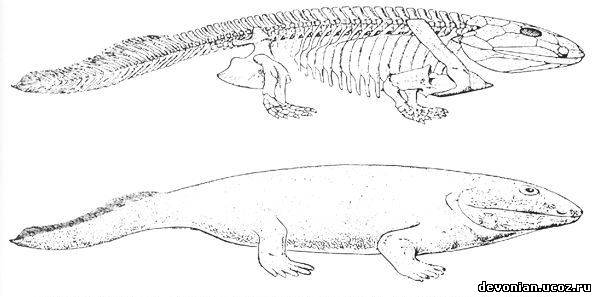
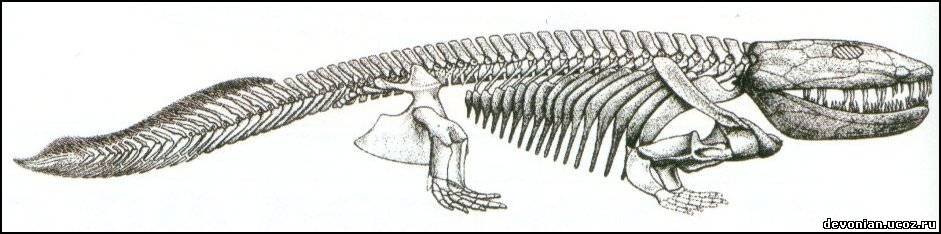
Шведские и британские палеонтологи из Университета города Упсала (Uppsala universitet) и Кембриджского университета утверждают, что загадочная "четвероногая рыба" ихтиостега - это вовсе не недостающее эволюционное звено между морскими и наземными животными, а просто один из неудавшихся природных "экспериментов", то есть эволюционный тупик. Доказательства приводятся в статье, опубликованной в последнем выпуске журнала Nature. Ихтиостега (Ichthyostega) обитала на территории нынешней Гренландии 360-410 миллионов лет назад - во времена девонского периода (поздний девон). Будучи одним из самых древних позвоночных нашей планеты, примитивнейшим родом земноводных, ихтиостега начиная с 1930-х годов (с тех пор, как был обнаружен первый почти полностью сохранившийся скелет этого ныне вымершего животного) претендовала на статус "отсутствующего звена" между рыбами и обитателями суши. Теперь шведско-британская группа исследователей представила новую реконструкцию этого классического животного, рисующую радикально иную картину форм его тела и образа жизни. Нужно заметить, что однозначная интерпретация окаменелостей этого существа всегда вызывала определенные трудности. Даже при том, что был собран почти целый скелет ихтиостеги, одной-единственной окаменелости, которая показывала бы животное целиком, все же нет. Вместо этого приходится собирать своеобразную головоломку, пользуясь информацией, полученной в ходе изучения нескольких различных окаменелостей. Первоначально это было проделано в 1950-х гг. профессором Эриком Ярвиком (Erik Jarvik) из Музея естествознания в Стокгольме, который разглядел в древнем животном крокодилоподобное крупное туловище, четыре крепкие лапы и примитивный позвоночник. Однако нынешние ученые с подобной интерпретацией не соглашаются. Пятилетние исследования привели к выводу о том, что строение позвоночника было совсем иным, туловище сформировано совсем не так, а задние конечности вообще можно сравнивать скорее с тюленьими ластами, чем с лапами наземных животных. Новая реконструкция снабжает ихтиостегу, общая длина которой достигала одного метра, хребтом, который удивительно близок для такого древнего наземного животного к позвоночнику современных млекопитающих. Все это означает, что для ихтиостеги был характерен очень необычный способ передвижения - ее тело как бы скользило вбок. Скорее всего, это существо жило ловлей рыб и примитивных сухопутных предков нынешних саламандр и ящериц, во время засухи спасаясь в болотах. Эти выводы во многом справедливы и для другой, еще более примитивной четвероногой рыбы из Гренландии, также жившей в девонский период, - акантостеги (Acanthostega, что в переводе означает "колючебронированная"). Однако, в отличие от акантостеги, ихтиостега обладала более крупным туловищем с ребрами, укрепленными наподобие черепицы, что делало верх ее тела полностью жестким, а ее задние конечности, скорее всего, были лучше приспособлены к сгибанию в вертикальной плоскости. "Ихтиостега, вероятно, перемещалась по земле довольно неуклюже, приподнимая верхнюю часть своего тела и перебирая передними конечностями при одновременном подтаскивании вперед своих задних плавников . Возможно также, что подобный способ передвижения комбинировался с изгибом спинного хребта дугой в вертикальной плоскости, позволяя ей двигаться вперед подобно какой-то гигантской гусенице ", - говорит профессор Пэр Альберг (Per Ahlberg) из Университета Упсалы (две различные манеры передвижения варьировались в зависимости от того, насколько быстро ихтиостеге требовалось двигаться). Он уверен, что ихтиостега - это вовсе не "недостающее эволюционное звено", а скорее один из нескольких недолгих эволюционных экспериментов с различными физическими формами и принципами передвижения в течение переходного периода от моря к суше. Альберг заявляет также, что более примитивная и менее "земная" на первый взгляд акантостега, кажется, расположена ближе к "основной линии" развития земноводных: "Ихтиостегу можно считать своего рода аналогом неандертальского человека, который расположен близко к нашей родословной линии и до некоторой степени может восприниматься в качестве одного из наших предшественников, однако в действительности это совершенно отдельная ветвь эволюции, закончившаяся тупиком, это вовсе не наш предок".
Перевел Максим Борисов
Устаревшие иллюстрации ихтиостеги

 (поздняя реконструкция)
Новая реконструкция (правильная)
Подвижность объёмных суставов конечностей у раннего тетрапода Ichthyostega
Происхождение тетраподов и переход от плавания к ходьбе – один из главных шагов
эволюции и многообразия наземных позвоночных. За время этого перехода модификации
конечностей - в особенности суставов и структур, направляющих движение, - фундаментально
изменили способы передвижения тетраподов1–4. Тем не менее, об анатомии конечностей ранних
тетраподов и способах их передвижения известно немного, а ведь это был очень критический
этап эволюции позвоночных. В данном исследовании мы представляем объёмную
реконструкцию знаменитого девонского тетрапода – Ichthyostega, а также качественный и
количественный анализ степени подвижности её конечностей. Мы показываем, что Ichthyostega
ещё не обладала типичными для тетраподов локомоторными навыками, такими как боковая
последовательная ходьба. В частности, она ещё не могла совершать вращательных движений
конечностями в достаточной степени, чтобы поднять тело над землей и поочередно шагать. Если
учесть, что способность к вращению вокруг продольной оси была присуща еще плавникам
тетраподоморфных рыб, то можно предположить, что ранние тетраподы прошли стадию
ограниченной подвижности плечевых и бедренных суставов. Или что ихтиостега была в этом
отношении уникальной. Мы пришли к выводу, что ранние тетраподы, имевшие такую же
морфологию скелета и подвижность конечностей, как у Ichthyostega, едва ли могли оставить
недавно описанные следовые дорожки из среднего девона.
Ранние тетраподы и их предки-рыбы ранее интерпретировались, как животные, способные
передвигаться по суше, опираясь на плавники или ноги. Впрочем, находки окаменелостей, сделанные
в 90-ых годах, показали, что первые позвоночные с ногами вели преимущественно водный образ
жизни, и что конечности развились еще до способности «ходить» по земле.
Еще позже была разработана версия о том, что локомоция с помощью задних конечностей впервые
появилась у лопастепёрых рыб еще задолго до появления не только передвижения по суше, но и самих
конечностей с пальцами. Эта модель предполагает то, что развитие таза и крестца с крепящимися к
ним мышцами не было напрямую вызвано взаимодействием плавника\ноги с субстратом.
Высказывались и предположения о том, что некоторые ранние тетраподы способом движения
походили скорее на тюленей, чем на саламандр. Таким образом, до сих пор ведутся споры о том, когда
же, как и от каких предшественников произошли способы передвижения современных тетраподов, и
вопрос с выходом на сушу остается неясным.
Чтобы прояснить вопросы эволюции локомоции ранних тетраподов, мы создали
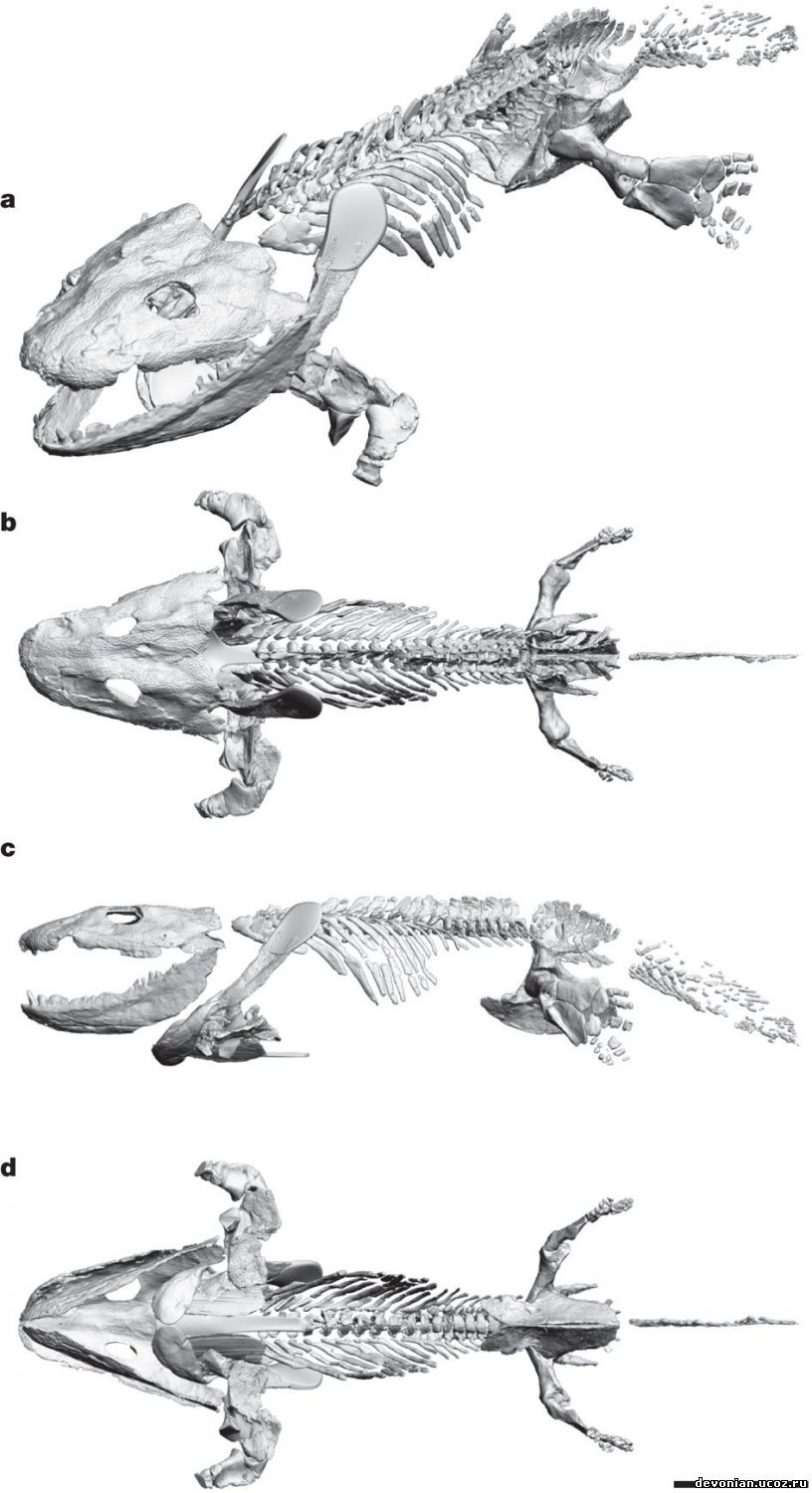
компьютерную модель сустава одного из наиболее известных девонских тетраподов - ихтиостеги. Для
этого мы использовали микротомографию, чтобы отсканировать наиболее сохранные образцы
ихтиостеги, получили трехмерную модель скелета (рис.1), и определили максимальный предел
свободы плечевого, локтевого, бедренного и коленного суставов в трех ортогональных плоскостях
движения. Чтобы интерпретировать степень подвижности суставов в контексте локомоции, мы
сравнили данные ихтиостеги с пятью морфологически и филогенетически разными современными
аналогами с разной морфологией суставов и разной манерой движения. Среди них были: саламандра
Ambystoma tigrinum, крокодил Crocodylus niloticus, утконос Ornithorhynchus anatinus, тюлень
Halichoerus grypus и выдра Lutra vulgaris. Кроме того, мы проводили анатомические вскрытия, чтобы
установить влияние наличия или отсутствия мягких тканей на подвижность суставов.
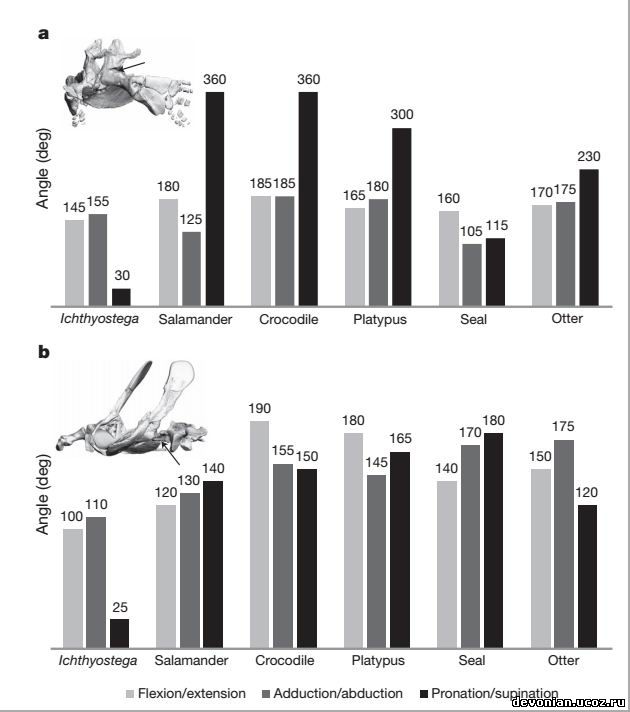
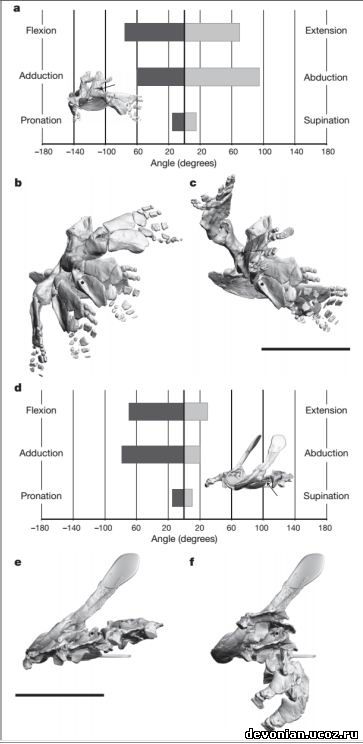
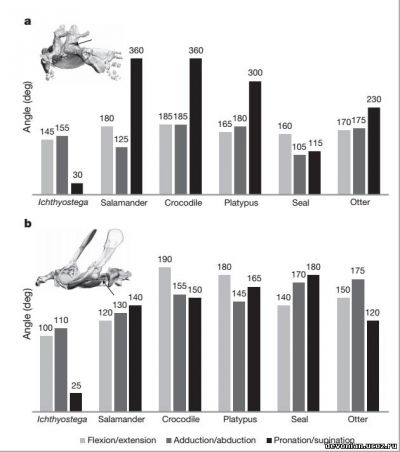
Как выяснилось, бедренный сустав ихтиостеги имел степень подвижности
(сгибание\разгибание, приведение\отведение), сравнимую с таковой современных тетраподов.
Впрочем, в отличие от современных тетраподов, бедренный сустав ихтиостеги имел минимальный
угол вращения вокруг продольной оси (пронация\супинация). Если отобразить степень подвижности
как позитивные и негативные угловые движения (рис. 3а), то бедренный сустав ихтиостеги
демонстрирует достаточную степень подвижности, приближающуюся к таковой у современных
тетраподов (см. Таб.5), и даже с большей амплитудой отведения. Что же касается коленного сустава,
Ichthyostega демонстрирует очень ограниченную подвижность в сгибании/разгибании (см. Таб.6), но
для двух других плоскостей движений не было обнаружено никаких видимых отличий, вероятно, по
причине того, что подвижность колена ограничивали мягкие ткани, как и у множества других
тетраподов, а не какие-либо остеологические препятствия.
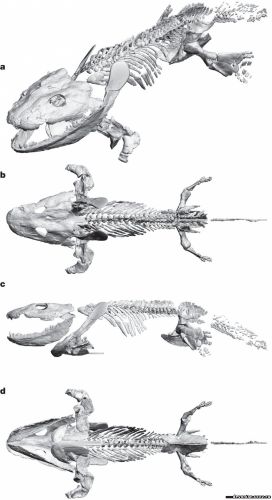
Бедренный сустав ихтиостеги образует мыщелкообразное сочленение с сильно увеличенными
дорсальным и вентральным костными выступами, окружающими вертлужную впадину, удлиненную
в передневентрально-заднедорсальном направлении. Кроме того, головка бедра имеет
бумерангоподобную форму с большой вентральной межвертельной ямкой, смыкающейся с костным
выступом вертлужной впадины, образуя ограничивающий механизм. Такой тип анатомии бедра
позволяет ему поворачиваться вокруг первичной и вторичной осей впадины, но не позволяет больших
вращательных движений вдоль продольной оси. Поскольку первичная ось вертлужной впадины (а
также головки бедра) наклонены вперед, задняя конечность могла двигаться лишь под углом 45
градусов к главной горизонтальной оси тела. Следовательно, задняя конечность двигалась по дуге в
передневентрально-заднедорсальном направлении во время приведения бедра и передневентрально -
заднедорсальном направлении во время отведения (рис. 3b,c).
Ограниченная подвижность вращения бедренного сустава по продольной оси вкупе с наклоном
сустава примерно на 45 градусов от горизонтали указывают на то, что подошвенная поверхность стопы
ихтиостеги не могла контактировать с субстратом. В частности, бедро не могло принимать
горизонтальную ориентацию, и стопа не могла направляться вперёд, то есть, принимать обычное для
наземных четвероногих положение. (рис. 5 и 6). Поэтому таз не мог подниматься над землей, а задние
конечности были нужны в первую очередь для плавания, тогда как на суше они выполняли более
пассивную или стабилизирующую функцию (примерно, как у тюленей). Если учесть развитие у
ихтиостеги широких веслообразных дистальных костей конечности и расширенных стоп, то
вырисовывается именно такая модель движения ее задних конечностей. В пользу этого говорит и
ограниченность сгибания\разгибания коленей - другого важного компонента ходьбы по земле.
Что касается передней конечности, то плечевой сустав ихтиостеги имеет наиболее
ограниченную амплитуду угловых движений во всех трех плоскостях (рис. 2b), и особенно
примечательно явное отсутствие вращения вокруг продольной оси (пронации\супинации). Если
перевести амплитуды движений в позитивные и негативные угловые движения, получится, что
плечевой сустав ихтиостеги был ограничен преимущественно негативными угловыми движениями, с
преобладанием сгибания и приведения (рис. 3d). В противоположность ихтиостеге, исследованные
современные тетраподы демонстрируют прекрасную подвижность плеча во всех трех плоскостях
движения (рис. 2b) и более равномерное распределение между позитивными и негативными угловыми
движениями (см. Таб.3).
Насколько можно судить по суставным поверхностям локтевой кости, локтевой сустав
ихтиостеги по подвижности был сравним с суставами современных тетраподов. Он был сравнительно
гибким во всех степенях свободы и демонстрирует схему, промежуточную между ползающими
животными и ходящими на выпрямленных конечностях (таб.4)
Ограничения движений в плечевом суставе у ихтиостеги, в первую очередь, обусловлены
переднезадним удлинением и дорсовентральным уплощением гленоидной ямки и головки плечевой
кости. Как и в случае с бедром, этот тип морфологии сустава образует кондилоидоподобное
сочленение, позволяющее сгибание\разгибание и приведение\отведение, но удерживающее от
вращения плеча вдоль продольной оси. Кроме того, гленоидная ямка ихтиостеги несколько скручена
вдоль своей основной оси: передняя часть направлена вентрально, а задняя - дорсально. Морфология
гленоидной ямки позволяет слегка смещать конечность в передневентрально-заднедорсальной
плоскости во время сгибания, а во время приведения она направляет конечность по наклонной
плоскости, заставляя плечо отклоняться кзади (рис. 3 е, f). В дополнение к форме сустава, плечевой
пояс ихтиостеги также имел большую костную выпуклость, окружающую переднедорсальную
границу гленоида, также ограничивающую разгибание и отведение.
Если учесть, что симметричная походка (например, последовательное латеральное шагание или
рысь) требуют больших углов разгибания и ротации, а также вращения плечевого пояса посредством
изгибов позвоночника (15, 16, 18-23), непохоже на то, чтобы ихтиостега могла передвигаться таким
образом. В действительности степень свободы плечевого сустава ихтиостеги в сочетании с
малоподвижными плечевым поясом и грудью (ввиду больших перекрывающих друг друга ребер), а
также с задними конечностями, неэффективными для передвижения по суше, указывают на то, что
наиболее вероятным способом передвижения по суше было ползанье с опорой на передние конечности
(наподобие как у илистого прыгуна). Кроме того, сильные мышцы, приводящие плечо, могли помогать
ихтиостеге поднимать голову над водой для дыхания или поимки добычи.
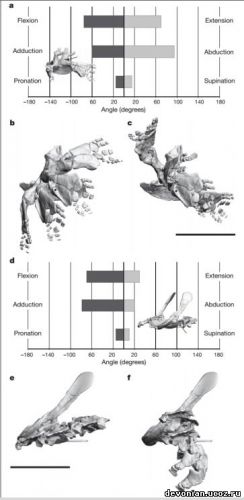
На основании исследования степеней подвижности суставов, а также морфологии ребер и
позвонков мы пришли к выводу, что ихтиостега не обладала «обычной» четвероногой походкой.
Способность поворачивать плечо и бедро вокруг продольной оси и способность к симметричной
походке, вероятно, развились у других видов ранних тетраподов. Известно, что подобный тип
морфологии плечевого и\или тазобедренного сустава присуствовала и у некоторых других ранних
тетраподов. Ограниченная (в основном, для вращения вокруг продольной оси) подвижность суставов
могла быть широко распространена среди них. Впрочем, определение наличия или отсутствия
симметричных походок у других ранних тетраподов требует создания таких же моделей конечностей
и осевого скелета. И все же способность к вращению вокруг продольной оси присуствовала у
плавников тетраподоморфных рыб, что свидетельствует о том, что некоторые тетраподы прошли через
фазу ограниченной подвижности плеча и бедра еще до приобретения способности к вращательным
движениям и моделей движения, характерных современным животным.
Кроме того, наши данные подтверждают то, что передние конечности тетраподов начала
использоваться для передвижения по суше раньше, чем задние, тогда как таз и задние конечности
служили преимущественно как дополнение к хвосту и использовались для перемещения под водой.
Для ходьбы задние конечности приспособились уже позднее (в противоречие источнику №14). Разная
функциональная роль передних и задних конечностей и более поздняя адаптация задних конечностей
к ходьбе также подтверждается меньшим развитием тазовых плавников у тетраподоморфных рыб по
сравнению с плечевыми, а также генетическими механизмами, контролирующими развитие
мускулатуры плавника и конечности. Также известно о заметном эволюционном расхождении в
развитии мускулатуры грудных и брюшных плавников у костных рыб.
В дополнение можно предположить, что ихтиостега еще не имела тазовой мускулатуры,
нужной для опоры на задние конечности, и на более поздних этапах эволюции «ходильные» группы
мышц стали постепенно преобладать над «плавательными».
Могла ли ихтиостега оставить следы вроде тех, что были описаны в среднем девоне? Все
доступные данные об анатомии суставов и осевого скелета доказывают, что подобные ихтиостеге
животные не могли оставить поочередных отпечатков ног. В частности, ранние тетраподы не могли в
достаточной степени совершать вращательные движения конечностей (и, возможно, делать
латеральные сгибания позвоночника), чтобы приподниматься над землей и поочередно шагать.
Возможно, следы оставил какой-то неизвестный вид тетрапода (или известный, но с несохранившимся
посткраниальным материалом) с иным строением суставов и аксиальной анатомией. Современные
научные данные пока не дают ответа на этот вопрос. Данное исследование говорит в пользу того, что
животные, подобные ихтиостеге, могли оставлять только синхронные отпечатки передних
конечностей.
Перевод на русский язык выполнил Сергей Паладий
|