Палеоэкология Dunkleosteus terrelli, автор Роберт Карр (Университет Конкордия, Чикаго).
Введение.
Наше понимание палеоэкологии позднедевонских ихтиофаун ограничено из-за того, что слишком мало фаун сохранилось во всем своем разнообразии, чтобы быть проанализированными. Кливлендская фауна - одна из самых известных и включает 66 видов (Hlavin, 1976; Denison,1979; Williams, 1990; Carr, 1996; Carr and Jackson, 2010), в том числе акул (33 вида), плакодерм (28) и костных рыб (5).
Палеоэкологические интерпретации фауны исторически основывались на сравнением с похожими видами из других фаун. Например, Coccosteus (Miles and Westoll, 1968), часто использовался как аналог для интерпретации других артродир (в т.ч. Heintz, 1932,p. 202, интерпретировал дунклеостея, сравнивая с коккостеем). Такие аналогии часто приводили к сравнению видов из разных периодов и несхожих фаунистических ансамблей.
Hlavin (1976) исследовал седиментологическую модель отложений в дельте Кэтскилла и черных сланцах (конкретно - Кливлендский компонент сланцев Огайо, а также другие сланцы этого региона и их биостратиграфические связи), и его фаунистический анализ Кливлендских сланцев до сих пор является самым актуальным. Однако он не учел влияния того, что образ жизни рыб мог повлиять на залегание их окаменелостей.
Williams (1990) провел первый сравнительный анализ пищевых цепочек Кливлендской фауны. В основном, он фокусировался на хрящевых рыбах ввиду их необычной сохранности - вплоть до содержимого желудков (описано 53 образца с годным к опознанию содержимым желудков - Williams, 1990, p. 280). Для нехрящевых рыб таких случаев нашлось всего несколько: акулий позвоночник в области рта артродиры; останки артродиры и акулы, предполагающие то, что это были хищник и жертва; останки палеонискидов в желудках акул и артродир(Williams, 1990, p. 286–287).
Несмотря на огромное количество работ по седиментологической истории Аппалачского бассейна (в т.ч. Woodrow и Sevon, 1985) и долгую историю палеонтологии позвоночных в этом регионе, палеоэкологии данной ихтиофауны было уделено мало внимания. Плакодермы, самая примечательная группа девонских рыб, составляющая наибольшую часть экспонатов в местных коллекциях, в палеоэкологическом отношении оказалась самой малоизученной.
Как видно из вышеприведенных примеров, палеоэкология палеозойских рыб основывается на таких фактах, как содержимое желудков, форма тела, отчасти - сам характер окружающих отложений. О содержимом желудков плакодерм известно мало, к примеру, Dennis и Miles (1981) а также Miles и Westoll (1968, p. 462) обнаруживали содержимое желудков у Incisoscutum и Coccosteus. Морфология челюстей может подсказать, какой была пищевая стратегия - дурофагия или ихтиофагия (Denison, 1978, p. 17). И, тем не менее, определить пищевые стратегии малоспециализированных форм или форм, не имеющих современных аналогов, остается трудным или даже невозможным без достаточного количества палеонтологических доказательств.
Lindsey (1978) и Webb (1982) использовали форму тела для интерпретации манеры движения современных рыб. Для плакодерм такой подход имеет ограниченное применение - ведь они делятся лишь на два основных типа формы тела: сплющенные латерально и сплющенные дорсовентрально. Miles (1969,p. 129; см. также Moy-Thomas и Miles, 1971, p. 175) предполагали, что латерально сплющенные артродиры вели нектонный образ жизни, тогда как некоторые другие авторы (Stensio ̈ , 1963, p. 13; Miles,1969, p. 129; Moy-Thomas and Miles, 1971, p. 185, 197–198) интерпретировали дорсовентрально сплющенные формы (например, ренаниды) как придонные. Между этими крайними вариантами есть множество видов, интерпретируемых как преимущественно придонные(Stensio ̈ , 1963, p. 13) или околодонные(Moy-Thomas and Miles, 1971, p. 197)
Наконец, анализ пород, в которых была найдена окаменелость, также может сообщить дополнительную информацию о среде, в которой обитала рыба. Трудность заключается в том, что ее останки могли посмертно попасть в среду, отличную от той, где она обитала.
Ни по одному из всех этих подходов полную картину палеоэкологии плакодерм восстановить невозможно, однако предпринимались попытки сделать их обобщенный анализ. Идеальная и полная сохранность организма, вплоть до мягких тканей и содержимого желудка - исключительно редкая удача. Такие сохранные образцы девонского периода известны по Кливлендской фауне (хрящевые рыбы в карбонатных конкрециях), (Williams, 1990). Что касается плакодерм, то наиболее сохранный материал был найден в формации Хунсрюкшифер (нижний эмский ярус Германии) и в формации Гого (фаменнский ярус, Западная Австралия). Лагерштеттские образцы тоже имеют хорошую сохранность - но неинформативны на счет палеоэкологии. Таким образом, лишь комплексный и всеобъемлющий подход, включающий анализ седиментологических, геохимический и тафономический механизмов, может предоставить достаточное количество информации об эволюции и палеоэкологии (Tasch, 1965; Elder и Smith, 1988).
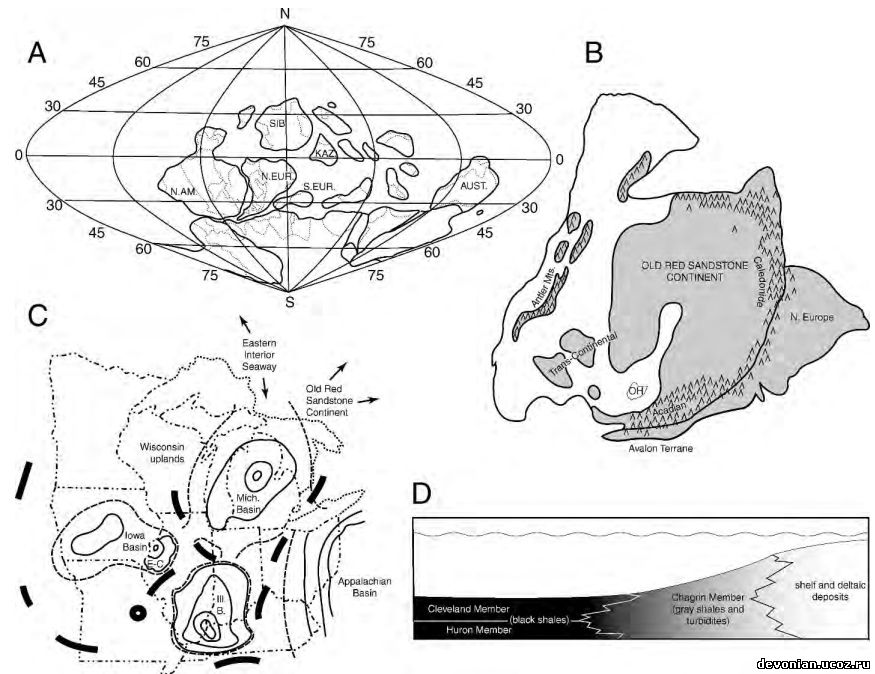
Дельта Кэтскилла и окружающий ее бассейн (Аппалачский бассейн, рис 1С) предоставляет уникальную возможность проанализировать палеоэкологию рыб этого региона. Черные сланцы(рис 1D) представляют собой осадки аноксической среды, вероятно, негостерпиимной для бентических организмов, и существовать там могла лишь незначительная фауна. Окаменелости рыб из этих сланцев представляют собой и рыб, обитавших в этом регионе, и останки, занесенные туда посмертно. (Brett и Baird, 1993, p. 254)
Исторически принято рассматривать большинство плакодерм как облигатные бентосные организмы, поднимавшиеся в толщу воды лишь для кормления(Stensio ̈ , 1963, p. 13; Moy-Thomas and Miles, 1971, p. 197;Denison, 1978, p. 17), указывая на о, что наличие их останков в малонаселенных глубоководных частях бассейна обусловлено вторичным накоплением трупов, происходящих из более мелководных и аэрированных частей бассейна.
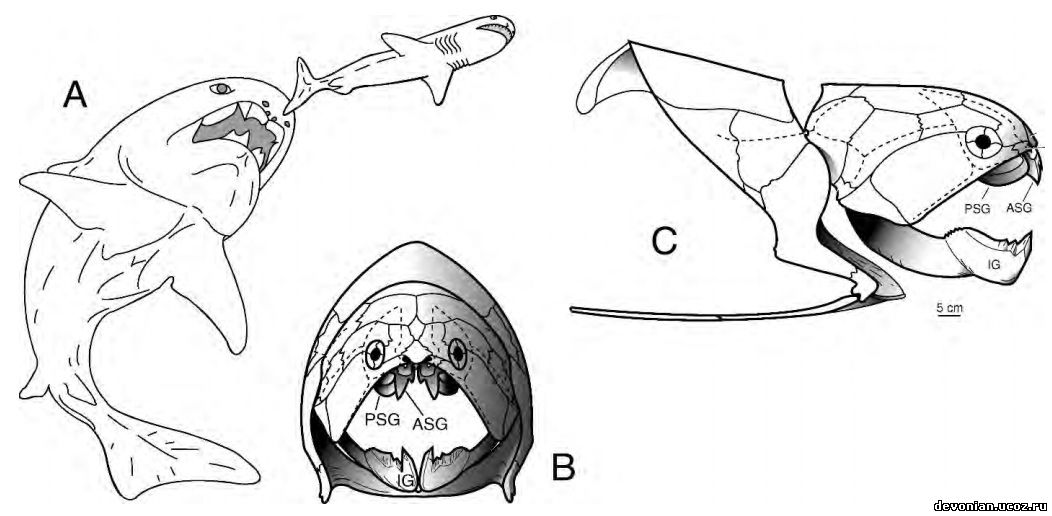
Чтобы подтвердить эту гипотезу на примере одного вида, данное исследование рассматривает распространения по Аппалачскому бассейну челюстных элементов Dunkleosteus terrelli (Newberry, 1873) Dunkleosteus terrelli(Рис. 2A–C) - крупная артродира (4,5-6 м в длину), обнаруженная в Аппалачском бассейне. Тело имеет овальное сечение, хорошо развит остеодермальный экзоскелет (Рис. 2B, C), отдельные пластины достигают толщины в 5-7,5 см. Челюстные элементы (IG,ASG, PSG, Рис. 2B, C) этого вида в процессе разложения отделялись первыми (см.Scha ̈ fer,1972, p. 49–91, и Elder, 1985, касательно процесса разложения туш) и, как следовало бы ожидать, должны находиться недалеко от места смерти животного (возможность вторичного перемещения рассматривается ниже). Географическое распространение челюстных элементов Dunkleosteus terrelli не подтверждает гипотезу о том, что трупы этой артродиры долго дрейфовали по акватории.
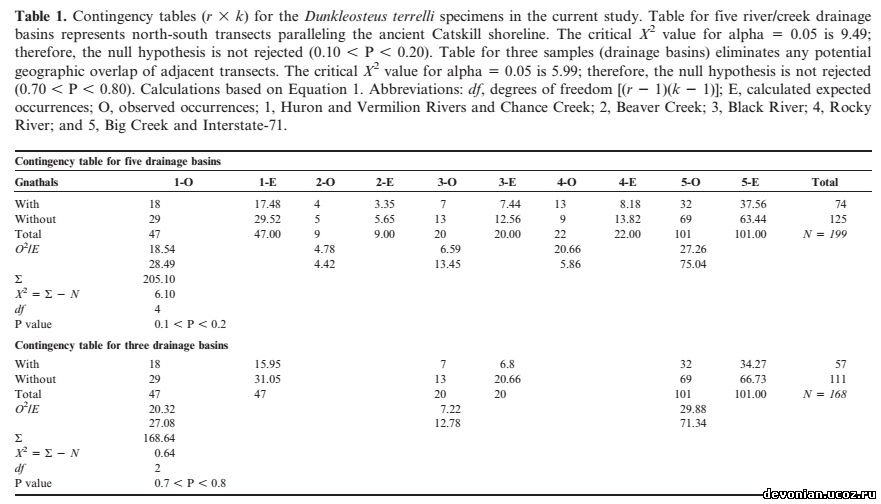
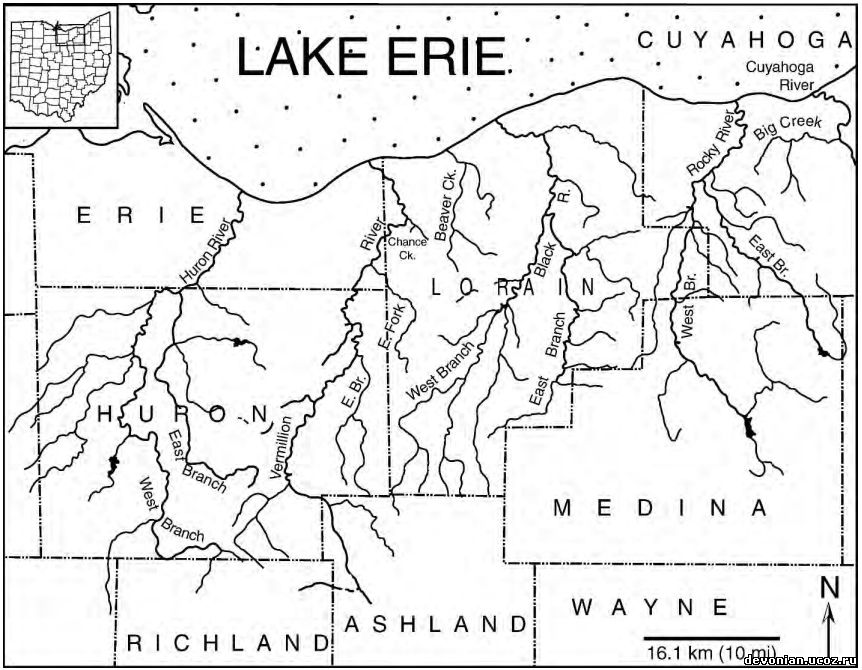
Образцы Dunkleosteus terrelli из Кливленда (верхний фаменнский ярус, конодонтная зона - между поздней expansa и ранней praesulcata, Zagger, 1995), Огайо, использованные для данного исследования, находятся в собрании Кливлендского музея естественной истории. Из Музея палеонтологии позвоночных были получены подробные сведения о местах находок этих образцов. Раскопки велись в семи бассейнах рек и ручьев, протекающих с севера на юг, а также в раскопе Interstate-71 в северном Огайо возле Кливленда (Рис. 3). Эти местности были сгруппированы в пять систем, идущих по северному Огайо с севера на юг, параллельно древнему побережью Кэтскилла (с запада на восток: 1. Реки Гурон и Вермийон, Ченс Крик; 2. Бивер Крик; 3. Блэк ривер. 4. Рокки ривер; 5. Биг Крик и Interstate-71). Эти бассейны обнажают черные сланцы (Кливлендский компонент). Описанные образцы включают в себя окаменелости с челюстными элементами, без них и отдельные челюстные элементы. Ниже будет приведено их сравнительное распределение в разных местностях.
Сухой вес - это вес живой особи, взвешенной вне воды, тогда как мокрый вес - это ее вес под водой. Масса тела (мокрый вес) - важный фактор, влияющий на плавучесть (статическую или динамическую) активных пловцов, не менее важный и для бентосных видов, а также сильно влияющий на посмертное перемещение трупов. Для определения локомоторных паттернов Dunkleosteus terrelli, его возможного сухого и мокрого веса использовалось сравнение с современными акулами западной Атлантики и Тихого океана. Всего было использовано 59 образцов 18-и видов с разной длиной и массой (Bigelow и Schroeder, 1948, все расчеты длины-массы основаны на их анализе). Bigelow и Schroeder (1948) предложили фактор пересчёта для сухого и мокрого веса акулы (5.5% для непостоянно плавающих и 2.6% для постоянно плавающих), который был применен для Dunkleosteus terrelli. Так как у акул отсутствуют кости, пересчёт получился приблизительным; тем не менее, он помог определить возможность погружения туши в донные осадки и плавучесть рыбы (статическую и динамическую).
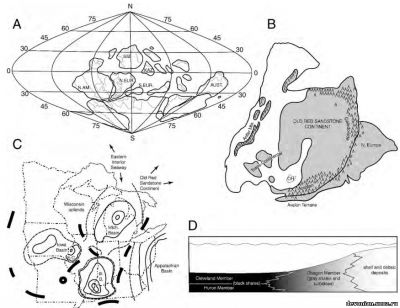
Рис. 1. А., палеогеографическая реконструкция с палеокоординатами на основа среднедевонской карты Streel и др., 2000. В. Детальная реконструкция североамериканского и североевропейского блоков по Ettensohn (1985, рис. 2). Отмечено расположение Огайо (ОН), предположительная суша затемнена, отмечены горы. C. Схема западного Аппалачского бассейна в Огайо и других региональных бассейнов (по Elliot и др., 2000, рис. 2). Отмечены границы штатов (штрихпунктир с двумя точками), Великие озера (точечный пунктир), контуры осадочных бассейнов (сплошные тонкие линии), гипотетические границы бассейнов (штриховой пунктир), региональные географические высоты (сплошные толстые линии). D. Схематический срез пересечения известняков Огайо (Кливлендский и Гуронский компоненты) и осадков береговой линии.
Обсуждение
Интерпретация палеоэкологии на основе разрозненных окаменелостей требует многочисленных предосторожностей (, Elder, 1985; Elder andSmith, 1988; Allison et al., 1991), но не теряет от этого своей важности. Элдер и Смит (1988) предложили метод интерпретации экологии рыб на основе тафономических данных. Основываясь на принципах теории информации (Tasch, 1965), они исследовали тафономию озерных рыб, и пришли к выводу, что тафономические процессы сами по себе дают полезную информацию. Во время тафономических процессов информация об организме теряется, однако сама степень тафономических изменений и их характер сообщают нам о физическом и химическом окружении организма, а также о его посмертной истории. А более детальное изучение тафономии позволяет судить не только об условиях захоронения организма, но и о его жизни. В отличие от лабораторных экспериментов, в которых рассматриваются индивидуальные вариации, исторические события требуют тщательного анализа всех тафономических вариаций. Каменная летопись плакодерм скупа и дает ограниченные данные об образе жизни и истории.
В Аппалачском бассейне окаменелости Dunkleosteus terrelli - самые многочисленные из всех позвоночных позднего девона. Это один из самых изученных представителей данной фауны (Heintz, 1932) и легко распознается благодаря большим размерам и уникальной остеологии, непохожей на таковую у других крупных представителей фауны (например, тонкие пластины и иная морфология титанихтиса легко отличают его от дунклеостея кливлендской фауны) Размеры особей Dunkleosteus terrelli варьируют от 25 см до 6 м, известны даже еще меньшие образцы, чья видовая принадлежность, впрочем, под вопросом. Таким образом, окаменелости представляют собой разные возрастные группы.
Материалы Dunkleosteus terrelli, хранящиеся в Кливлендском музее естественной истории, собирались на протяжении десятилетий. Питер А. Бангарт (Hlavin, 1976) собрал с 1923-го по 1946-ой множество материала в районе округов Сайяхога и Лорейн в северном Огайо (Рис 3), тщательно и щепетильно реконструируя из разрозненных фрагментов отдельные особи - даже если находки разных фрагментов разделяло несколько лет. Эта коллекция, вероятно, представляет собой наилучшее для статистического анализа собрание палеозойских образцов, с наименьшей погрешностью при проведении раскопок.
И хотя вопрос о происхождении аноксических бассейнов (Hoover, 1960, p. 31; Degens et al., 1986; Wignall, 1994;Schieber, 1998) остается дискуссионным, ряд факторов является для разных гипотез общим - в частности, касательно формирования черных сланцев, таких как в Кливлендском компоненте. В их числе можно назвать:
1) минимум или отсутствие доступного кислорода(Hoover, 1960; Heckel, 1972; Degens and Ross, 1974;Brumsack and Thurow, 1986; Wignall, 1994; Allison et al., 1995);
2) наличие гидроген-сульфида - возможного токсина(Hoover, 1960, p. 34; Heckel, 1972; Jannasch et al., 1974);
3) нестабильный субстрат, состоящий из тонких осадков высокой пористости(Keller, 1974);
4) мало освещаемая толща воды;
5) ограниченное количество или отсутствие окаменевших следов или признаков биотурбации
Сочетанный эффект первых трех факторов очевидно влияет на формирование бентических сообществ. Наличие аноксических вод признается всеми гипотезами, однако происхождении аноксии трактуют по-разному (к примеру, от вздымания осадков в толщу воды).
Опубликованные описания бентосных сообществ черных сланцев разнятся от полного отсутствия бентических организмов(Hoover, 1960, p. 32, 42; Conantand Swanson, 1961, p. 56–62; Heckel, 1972; Hlavin, 1976; Allisonet al., 1995, p. 100) до неразнообразных сообществ, приспособившихся переносить особенности своей среды обитания (аноксия или дизоксия, постоянная или перемежающаяся, гидроген сульфид, глубина ниже фотической зоны, нестабильный субстрат - по Hannibal et al., 2005). Детальные геохимические (о степени пиритизации) и седиментологические данные по Кливлендскому компоненту ограничены, за исключением недавних исследований латерально эквивалентных черных сланцев, других черных сланцев того же периода, или современных аналогов.
Современные бедные кислородом зоны на границе воды и осадка известны по множеству регионов, отличающихся высокой продуктивностью: к примеру, северо-запад Индийского океана (Degens et al., 1986), Калифорнийский залив (Brumsack and Thurow, 1986). Однако есть несколько примеров стагнантных внутренних морей: например, Черное море (Degens and Ross, 1974) или норвежские фьорды (Brumsack and Thurow, 1986). Это возможные аналоги древних аноксических систем и Аппалачского бассейна. Черное море характеризуется пониженной степенью седиментации (разной в разных частях бассейна) и высокой пористостью осадков, которые могут содержать до 71% (или более) всего объема воды (Keller, 1974). Седиментация происходит в бескислородной толще воды, насыщенной сероводородом. Несколько верхних миллиметров осадков не сохраняют даже остатков микроорганизмов. Следующие 30-60 см имеют консистенцию кашистой суспензии (Keller, 1974, p. 333). Прочность при скалывании не увеличивается вплоть до глубины примерно в 140 см (Keller, 1974,fig. 2). Под воздействием давления аноксической воды и присутствия сероводорода происходит образование и накопление сульфидов железа (Berner, 1974; Rozanov et al., 1974). Ни Черное море, ни фьорды, впрочем, не являются явными копиями больших внутриконтинентальных бассейнов девона; однако осадки, накапливающиеся в них, потенциально могут стать черными сланцами.
Кливлендский компонент (Nelson, 1955; Hoover, 1960; Mausser, 1982) - типичный представитель черных сланцев с высоким содержанием органики, сформировавшихся в бескислородной среде. От свежего излома таких сланцев исходит нефтяной запах из-за высоко содержания органики (Hoover, 1960, p. 23). Это композиция из тонких осадков с низким содержанием или отсутствием карбонатов в глубине сланца (Mausser,1982, p. 86; хотя и присутствуют непостоянные конические и карбонатные конкреции), что подразумевает нестойкий пористый субстрат, имевший место в момент залегания осадков (он мог в тот момент содержать до 70-80% воды Schieber, personal communication, 2004). Наличие диссеминированного пирита(Mausser, 1982) во всей формации указывает на присутствие в момент залегания сульфида водорода.
В северном Огайо породы верхнедевонского Аппалачского бассейна обнажаются в ряде бассейнов рек, текущих с севера на юг и впадающих в озеро Эри (рис.3). Эти бассейны представляют собой серию трансект, приблизительно параллельных древней дельте Кэтскилла (рис 1В, С), и предоставили ряд образцов черных аппалачских сланцев из разных частей бассейна от проксимальной до дистальной.
Кливлендский компонент представляет собой дистальный элемент поверхности бассейна, фактически эквивалентный восточному склону отложений Чагринского компонента (рис 1D). Эта прибрежная поверхность повторяет паттерн Аппалачского бассейна, как отмечали Baird and Brett (1986).
Анализ палеогеографического распространения Dunkleosteus terrelli позволяет сделать две потенциальные гипотезы: (H1) D. terrelli жил, умирал и сохранялся внутри бассейна; (H2) D. terrelli жил и умирал в другом месте, однако заносился в бассейн, где и сохранялся. Палеогеография D. terrelli отражена в природе и распределении его останков по Кливлендскому компоненту. Гипотеза 1 предполагает то, что останки этого вида могли сохраняться как интактными (без посмертного распада), так и расчлененными (если расчленение происходило в процессе перемещения туши - то перенос туш в глубокие части бассейна с востока должен быть исключен). Гипотеза 2 подразумевает, что останки D. terrelli заносились в глубокий бассейн извне. В пользу второй говорит гипотеза о расстояниях посмертного перемещения (т.е., более целые образцы должны обнаруживаться ближе к месту обитания, чем разрозненные).
Останки Dunkleosteus terrelli в Кливлендском компоненте обычно в той или иной степени расчленены, и та или иная часть пластин у них отсутствует (только 5,1% известных образцов сохранили более 25% дермальных и перихондральных костей). Elder (1985) и Elder и Smith (1988) описали три возможных причины расчленения и утраты органических останков в водной среде: дрейф, воздействие падальщиков и перенос течениями (см. также Allison и др., 1991).
В дистальной части Аппалачского бассейна два последних механизма имеют малое влияние или вообще не имеют его. Следует учитывать аноксию и условия формирования черных сланцев: бескислородные условия (с присутствием сульфида водорода, по некоторым моделям) могли сделать дно непригодным для обитания макроскопических падальщиков; впрочем, это не отрицает пожирания туш падальщиками еще в толще воды. То, что дно было непригодным для жизни, подтверждается отсутствием или незначительным количеством следов передвижения и биотурбации. Нет свидетельств о наличии разнообразной макроскопической инфауны или эпифауны, а также признаков обрастания костей эпибионтами. Окаменелости акул в Кливлендском компоненте часто обнаруживаются целыми и с сохранившимися мягкими тканями(Williams, 1990). Эти окаменелости обнаруживались в зонах карбонатных конкреций в сланцах Биг Крик и Интерстейт-71. В противоположность отдельным сплющенным образцам, эти окаменелости легко просматриваются в профиль. Окаменелости акул, найденные вне зон конкреций в западных отложениях, обычно ассоциируются с коническими отложениями или встречаются отдельно внутри сланцев. Скелеты этих рыб состоят из перихондрально окостеневшего призматического хряща, очень уязвимого к механическим повреждениям и легко уничтожающегося падальщиками и течениями.
Степень зернистости осадков Кливлендского компонента предполагает сравнительно малоэнергетическую среду ((Nelson, 1955; Mausser, 1982) и отсутствие течений, достаточных для перемещения больших костей Dunkleosteus terrelli.Кроме того, отсутствуют седиментарные структуры, предполагающие наличие в Кливлендском компоненте сильных придонных течений. Пересеивание осадков имело место, что доказывает наличие некоторых отдельных задержанных наносов (танатоценозы беспозвоночных - Hlavin, 1976; илистые наносы - J.Schieber, личная переписка, 2004); впрочем, пересеивание располагает скорее к накоплению костей и пелагических беспозвоночных, чем к их удалению. Крупные и сравнительно тяжелые кости Dunkleosteus terrelli должны были предположительнее оставаться в этих накоплениях.
Посмертный дрейф представляется наиболее правдоподобным объяснением находкам разрозненных останков Dunkleosteus terrelli. Такой дрейф мог происходить и у рыб, обитавших непосредственно в бассейне, и у организмов, или приводить к занесению в бассейн организмов, живших вне его.
Общей чертой процесса разложения и у водных, и у наземных организмов является потеря фрагментов тела под воздействием падальщиков (микроскопических и макроскопических). Орофарингеальная полость под их воздействием может разрушиться и лишиться челюстных элементов. У телеостов и артродир (Рис. 2В, С) кости верхней и нижней челюстей, не имеющие кострных соединений с осевым скелетом, утеряны. В отличие от них, у млекопитающих, чьи верхние челюсти и предчелюстные кости прочно соединены с черепом, теряются только дентарии.
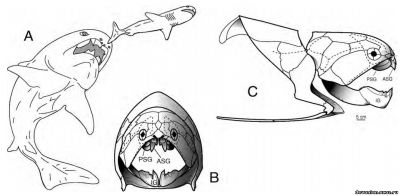
Рис 2. A, реконструкция живого Dunkleosteus terrelli, преследующего Cladoselache. По Carr (1995, рис. 17). B и C. Костный скелет D. terrelli, вид спереди и справа (по Heintz, 1932 и 1968). ASG 5 – передняя надчелюстная кость, IG 5 – подчелюстная кость, PSG 5 – задняя надчелюстная кость, пунктиром обозначены сенсорные каналы.
Работы Elder (1985) и других авторов (Scha¨fer, 1972; Hill, 1979; Smith and Elder, 1985; Elder and Smith, 1988) подтверждают, что челюстные элементы (у млекопитающих - дентарии) очень часто в процессе разложения отпадают рано, и наличие образцов с сохранными челюстными элементами объясняется сравнительно быстрым захоронением и недолгим воздействием предыдущих тафономических процессов (в том числе перемещения и падальщиков). В случае с Dunkleosteus terrelli в Аппалачском бассейне наличие отдельных челюстных пластин говорит в пользу того, что плавающие туши могли терять фрагменты сравнительно недалеко от места обитания.
Elder (1985), Smith и Elder (1985), и Elder и Smith (1988) обосновали важность глубины воды и её температуры для процесса флотации. Физические принципы, исходящие из газовых законов Шарля, Гей-Люссака и Бойля, отчётливо определяют плавательный потенциал организма, подвергающегося бактериальному разложению в границах заданных глубины и температуры. Это значит, что объем газов, накапливающихся в полостях тела и в тканях в процессе разложения и заставляющих труп плавать, прямо пропорционален температуре (климатический фактор) и обратно пропорционален глубине (фактор давления). Elder (1985) предположила, что ограниченная глубина дрейфа, обнаружившаяся у некоторых костистых рыб в его экспериментальной работе, не будет наблюдаться у крупных видов или при различной плотности. Она высказала предположение, что залегание трупа на глубине 10-и метров даже в тропических водах начисто исключает возможность его всплытия. В противоположность, Allison и др. (1991) высчитали плавательный потенциал у китов на глубине 1200 м. В отличие от костистых рыб в экспериментах Elder, туши китов имели примерно нейтральную плавучесть и с самого момента смерти дрейфовали, зависая в толще воды.
Аппалачский бассейн позднего девона находился в субтропиках (30–35 градусов южной широты), а значит, температура воды в нём была от умеренной до тёплой (17-18 градусов Цельсия) - она была рассчитана по изотопам кислорода в брахиоподах (Streel и др., 2000) Некоторые авторы по-разному оценивают его глубину - впрочем, все эти оценки хорошо вписываются в пределы, согласно Allison и др. (1991), необходимые для дрейфа:
61-91,4 м - по Nelson (1955);
30,5-45,7 м - по Hlavin (1976);
50-100 м - по Brett и др. (2003);
до 100 м - по Ferguson (1963).
Сравните с оценкой Schieber (1998) - 10-20 м в сланцах Чаттануги (штат Теннесси). Впрочем, сланцы Чаттануги характеризуются множеством задержанных наносов и эрозированных поверхностей.
Наличие останков D. terrelli в чёрных сланцах Аппалачского бассейна можно объяснить как посмертным накоплением организмов, непосредственно обитавших в этом бассейне, так и «дождём» из фрагментов, падающих с дрейфующих туш, заносимых в бассейн из богатых кислородом регионов. Результаты статистического анализа этих окаменелостей не подтвердили гипотезу о том, что D. terrelli ограничивался мелководной и аэрированной восточной частью бассейна и что его наличие в отдаленных осадках было результатом заноса туш в бассейн (таб. 1, Х2 5 6.1. d.f. 5 4, 0.10, P, 0.20).
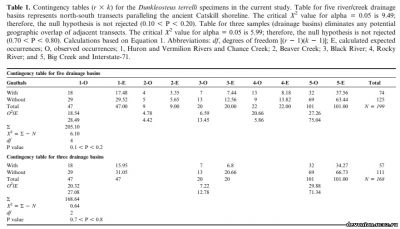
Дополнительный анализ непримыкающих срезов (nos. 1, 3, and 5) не позволяет и отвергнуть первоначальную гипотезу (X2 5 0.64, d.f. 5 2, 0.70 , P , 0.80). В её пользу говорит то, что D. terrelli случайным образом распространён по бассейну и скорее вел пелагический (и обитал и умирал на всей площади бассейна), чем сугубо придонный образ жизни(с привязкой к бентосу восточных аэрированных мелководий - напр., Чагринский компонент и более прибрежные регионы, рис 1 D).
Если приобщить наши знания о придонной экосистеме, связанной с чёрносланцевой формацией, получается, что D. terrelli должен был быть выносливым пловцом. Палеонтологические, седиментологические и геохимические свидетельства указывают на то, что придонная среда была негостеприимной; впрочем, даже если рыбы могли доныривать до дна бассейна, стабильность субстрата не позволила бы выдерживать их вес.
На данный момент оценок размера и веса Dunkleosteus terrelli немного (Heintz, 1932). Большое количество костей и, очевидно, большая масса тела натолкнула некоторых исследователей на мысль о том, что D. terrelli был бентической рыбой (в т.ч. Denison, 1978; впрочем, отрицается другими авторами; Heintz, 1932, пришел к выводу, что Dunkleosteus terrelli должен был быть активным пловцом и хищником). Сравнение Dunkleosteus terrelli (использовалась взрослая особь длиной в 4,6 м) с современными акулами (рис. 4) позволяет оценить сухой вес в 665 кг. Bigelow и Schroeder (1948) определили, что мокрый вес современной акулы составляет 5,5% от сухого веса (2,6% у активно плавающих форм). Используя эту оценку, мокрый вес D. terrelli можно определить как 36,6 кг (или 17,3 кг, если использовать процент для активных пловцов). Это оценка приблизительна, если учесть отсутствие у акул костей и влияние на плавучесть жира. Такой вес вполне достаточен для погружения D. terrelli на субстрат, что привело бы к интоксикации и забиванию жабер мелким илом (по Keller, 1974). Если D. terrelli обитал в глубокой части бассейна - то ему бы пришлось постоянно плавать, избегая отдыха на дне.
Вторая подсказка к интерпретации образа жизни плакодерм — их тип локомоции. Плакодермы, по всей видимости, обладали низким профилем и сравнительно слабым гетероцеркальным хвостом, поэтому можно предположить, что они плавали посредством синусоидальных волнообразных изгибаний тела (Thompson, 1971; угреобразная локомоция по Lindsey, 1978). Их сравнивают с современными макруридами или рыбами-химерами (Stensio, 1963), которые демонстрируют придонный образ жизни. И хотя угреобразный тип локомоции можно счесть сравнительно неэффективным способом плавания (если сравнить его скорость с другими способами плавания), это отнюдь не значит, что «угреобразные» пловцы не способны к длительному плаванию. Достаточно вспомнить маршруты миграции современных угрей (Anguilla anguilla), путешествующих из Северной Америки в Европу (McDowall, 1988), безошибочно находя свою родину. И хотя плакодермы, вероятно, никогда не развили совершенных типов локомоции, подобных таковым у современных костных рыб (например, у каранксов и тунцов, Lindsey, 1978), они развили ряд локомоторных адаптаций, усиливающих подъемную силу и уменьшающих трение (Carr, 1995).
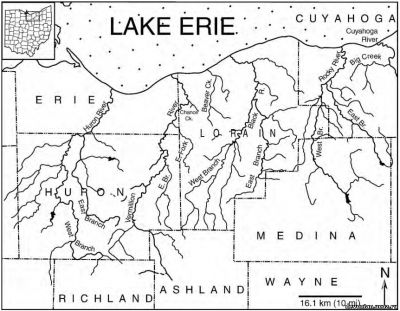
Рис. 3. Гидрологическая карта северного Огайо, в том числе дельты семи рек и ручьев, в которых были найдены все образцы, использовавшиеся в данном исследовании. В углу – карта штата Огайо, прямоугольником обведена исследованная территория. Крупным шрифтом обозначены округи, мелким – названия рек и ручьев. По данным департамента природных ресурсов штата Огайо, и федерального геологического бюро в штате Огайо, карты от 1985 и 1969 годов.
Заключение
Тафономические свидетельства не подтверждают версию о том, что Dunkleosteus terrelli был облигатным бентическим организмом. Статистический анализ распространения останков этого вида полностью отрицает гипотезу о сугубо донном образе его жизни. Dunkleosteus terrelli свободно плавал по Аппалачскому бассейну, и это объясняет его широкое распространение по Северной Америке (от Калифорнии до восточных штатов; если синонимизировать его с D. marsaisi (Lehman, 1954) из Марокко (Rucklin, 2002), то ареал расширится через океан Реикум до самой Лавруссии, рис. 1А). Эта версия находится в согласии с исследованием Carr (1995), описавшего развитие локомоторных специализаций у пахиостеоморфных артродир, а также затем была подтверждена распространением по бассейну сравнительно полных образцов и отдельных элементов, потерянных в результате разложения.
Аргументы против версии о свободноплавающих плакодермах — наличие у них тяжелой брони и угреобразного типа локомоции. Распространение целых и разрозненных останков Dunkleosteus terrelli в Аппалачском бассейне можно объяснить их посмертным дрейфом. Возможность посмертного дрейфа в данном бассейне, глубина которого отценивается в 30-100 метров, предполагает наличие у организма некоторой положительной плавучести (по Allison и др, 1991). Угреобразный способ плавания необязательно является неэффективным. Похоже, что вывод о придонном образе жизни основан на том, что исследователи выбирали для сравнения с плакодермами современные придонные формы, чем на настоящей корреляции между образом жизни и способом передвижения.
Выводы данного исследования троякие и станут основой для дальнейших работ над фауной Кливлендского компонента. Возникли следующие вопросы:
1) репродуктивная стратегия;
2) жизненный цикл;
3) статическая плавучесть Dunkleosteus terrelli.
Негостеприимное дно делает немногочисленными потенциальные места для нереста. На сегодняшний день неясно, был ли этот вид яйцеродящим или живородящим; в собрании Кливлендского музея естественной истории есть предполагаемые яйцевые капсулы данной рыбы (CMNH 8133–8136, 9461), что вызывает интересные вопросы. Нужна и дальнейшая работа по статистике возрастных групп D. terrelli в Аппалачском бассейне. И наконец, сам факт дезартикуляции, связанной с дрейфом туш, предполагает наличие у D. terrelli нейтральной плавучести. Elder (1985) отмечал, что на посмертную флотацию влияют физические факторы (глубина и температура); впрочем, Allison и др. (1991) продемонстрировали, что достаточная статическая плавучесть может наблюдаться на глубинах вплоть до 1200 метров. Наличие у плакодерм воздушных полостей не доказано (Denison, 1941, предположил их наличие у Bothriolepis, однако это предположение спорно, и, тем более, нельзя быть уверенным, что это было общей чертой всех плакодерм; Gardiner, 1984, утверждал, что воздушный пузырь — уникальное «изобретение» костных рыб). Тем не менее, наличие дезартикуляции, происходившей во время дрейфа трупов D. terrelli, уверенно говорит в пользу наличия какого-то механизма нейтральной плавучести у этого вида (возможно, за счет жира — как у хрящевых рыб). Дальнейшие работы по тафономическим паттернам и плавучести D. terrelli могут пролить свет на эти вопросы.
Перевел Паладий Сергей |